強化生物除磷系統中的生物化學原理
強化生物除磷系統中的生物化學原理
曹雪梅
(哈爾濱工業大學 市政環境工程學院 哈爾濱 150090)
摘要: 概括了國內外學者在研究EBPR系統生物化學原理方面提出的模型和觀點,并指出一些尚未解決的問題,提出EBPR的生物化學原理是一個很有價值的研究課題。
關鍵詞:強化生物除磷 PAO 醋酸鹽 還原力
The Biochemistry of EBPR System
Cao Xuemei
(School of Municipal & Environmental Engineering, Harbin Institute of Technology, Harbin 150090)
Abstract: This paper aims to generalize the models and views on the biochemistry of EBPR system that in-country researchers put forward , and point out some problems that haven’t been resolved, and suggest the biochemistry of EBPR system be a valuable research task.
Keywords: Enhanced biological phosphorus removal ,PAO, Acetate, Carbon source
為有效控制水體富營養化,迫切需要去除排放廢水中的植物性營養物——氮和磷。完全采用活性污泥系統進行微生物除磷的工藝被稱為強化生物除磷(EBPR)工藝。迄今為止,強化生物除磷工藝已經有四十幾年的歷史,EBPR工藝由三段Bardenpho或Phoredox工藝演變而來。現在廣泛使用的Johannesburg和改良的UCT工藝都是EBPR工藝的代表。EBPR工藝雖然具有較高的除磷效率,但是由于對強化生物除磷過程的了解甚少,因此污水廠的設計和運行條件不可能是EBPR所要求的最佳條件,進而導致污水廠的運行不穩定。因此有必要探討研究強化生物除磷的生物化學原理,從而設計出最優化的EBPR污水廠。筆者就近幾年國內外學者對EBPR生物化學原理的研究進行了總結。
1.反映EBPR生物量特征的化學變化
有關EBPR系統中生物量在厭氧/好氧循環期間發生的總的化學變化,許多文獻都有介紹。研究者普遍認為在厭氧區,一些細菌能快速吸收由化能有機營養菌發酵底物形成的短鏈揮發性脂肪酸VFA,VFA不支持細胞的生長,而是以聚β羥基鏈烷酸鹽(PHA)的形式貯存在細胞內。PHA的化學組成由底物決定:當底物為醋酸鹽時,合成聚β羥基丁酸鹽(PHB);而當底物為丙酸鹽時,主要產生聚β羥基己酸鹽。在厭氧階段,在吸收醋酸鹽的同時生物量中PHA的含量增加,細胞內的聚磷酸鹽含量下降,從生物量中釋放出的磷酸鹽使得上清液中磷酸鹽的含量上升。在接下來的好氧階段,生物量中PHA的含量下降,同時上清液中磷酸鹽的含量也下降,而生物量中聚磷酸鹽的含量上升,含磷量大于細胞干重的15%。如果從好氧反應器中取出一定量的生物量樣品放到顯微鏡下觀察,能檢測到染色的富含聚磷酸鹽的細胞,但檢測不到含PHA的細胞。另一方面,從厭氧反應器中取出生物量樣品,經過染色,放到顯微鏡下觀察到含PHA的細胞,卻沒發現含聚磷酸鹽的細胞。這些化學變化能反映EBPR生物量的特征。通常假設這些化學變化只由聚磷菌(PAO)來實現。盡管良好的強化生物除磷效果經常與生物量中出現的大團緊密的細胞簇有關,但是有關PAO細胞簇組織對EBPR的影響尚不清楚。
2.EBPR系統的生物化學原理
2.1以醋酸鹽作為唯一碳源時EBPR的代謝作用
一些經驗模型都試圖解釋EBPR的化學變化,這些模型主要是通過以醋酸鹽作為唯一碳源時得到的數據來分析研究生物群落的化學變化,如圖1所示[3,4]。所有模型都是建立在分析化學數據的基礎上,這些數據反映總的微生物群落的變化,但不能反映個別種群的變化。模型假設聚磷酸鹽的吸收與釋放,PHA的合成與再利用都是由一種PAO種群實現的,所有的細胞都有相同的代謝屬性。
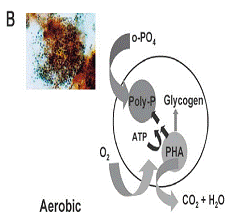
Fig. 1. A summary of the major features of the biochemical models for EBPR.[3,4]
這些模型對EBPR之所以能高效率除磷的解釋是:由于在厭氧/好氧循環的條件和底物缺乏的條件下,PAO能優先利用底物合成貯存在細胞內的化合物,從而成為優勢菌屬。因而,在厭氧條件下,PAO能快速吸收像醋酸鹽這樣的有機底物,以貯存的聚磷酸鹽為能量來源合成PHA,并釋放正磷酸鹽。在缺乏有機物的好氧區,微生物將體內貯存的PHA作為碳源和能源,以維持其生長,并吸收磷酸鹽合成聚磷酸鹽。因此,PAO在前置厭氧/好氧的條件下成為優勢菌屬是因為它們能在缺少碳源和能源的好氧條件下,利用厭氧條件下積累的PHA作為碳源和能源來維持其生長。一般認為在底物缺乏的生態系統中,PAO能更好的存活,這是因為PAO能快速吸收底物,貯存能量和有機物,其生長速率大于其他細菌。合成PHA需要還原力,各種模型所提出的還原力有所不同。Comeau[1]和Wentzel[2]等人認為合成PHA所需要的電子來源于TCA循環,但是Mino等人[3,5]證明合成PHA所需的電子是由PAO好氧合成的貯存在細胞內的糖原在厭氧條件下分解代謝產生的。目前,大多數學者都認為Mino模型是正確的。但是Pereira[6]和Hesselmann[7]等人認為只靠降解糖原是不能滿足PHA合成所需的還原力,還需要TCA循環提供部分電子。糖原通過控制PAO的氧化還原平衡而對EBPR起著關鍵性的作用,這一點對實際的EBPR系統允許PAO厭氧吸收和代謝不同種類的可生物降解的底物是必要的。某些事實表明在受沖擊負荷影響的條件下,糖原的可利用量而不是細胞內的聚磷酸鹽量,會最終限制細胞厭氧吸收底物的能力。
研究者圍繞PAO厭氧吸收底物和合成PHA過程的生物能量學展開大量爭論。早期的研究認為降解聚磷酸鹽釋放的ATP是PAO吸收底物,合成PHA,進行其他生物合成反應,維持細胞生長的唯一能量來源。但是現在也有人認為糖原的分解代謝能為合成PHA提供ATP,而其提供的可利用的能量的多少由糖原分解代謝的途徑決定。
利用核磁共振可以跟蹤顯示混合的生物群落在特定的且特殊的底物供應的條件下的代謝物的量,這種方法可以解釋化學數據所帶來的迷惑與矛盾。利用核磁共振,Pereria[6]等人說明了EBPR厭氧運行時生物量的TCA循環,并指出糖原是以ED(Entner—Doudoroff)方式降解而產生合成PHA所需的還原力。重要的一點是,醋酸鹽和糖原似乎都能被用來厭氧合成PHA,然后在好氧條件下PHA再被用來產生糖原。
Hesselman[7]等人證實了TCA循環可以作為PHA合成時還原力的部分來源且糖原的分解代謝是以ED方式進行的。他們在理論的基礎上特別指出,細胞不是以之前假設的主動運輸的方式吸收有機物,而是通過被動擴散或促進擴散來吸收醋酸鹽,即不需要消耗能量。而且,在厭氧釋磷代謝過程中最需要能量的一步反應,即吸收醋酸鹽的過程,并不需要醋酸鹽激酶的參與,相反卻利用了乙酰輔酶A。而在早期的模型中一直假定在反應過程中醋酸鹽激酶是很重要的。
能量來源對厭氧釋磷有重要的作用。早期模型提出的能量來源包括由聚磷酸鹽的降解而釋放出的大量焦磷酸鹽。焦磷酸鹽、磷酸烯醇丙酮酸鹽和AMP磷酸轉移酶都參與完成底物水平磷酸化,建立質子推動力。大量試驗證實厭氧釋磷量與醋酸鹽的吸收量的比值不是常數,此比值由混合液的pH值決定,pH值影響醋酸鹽運輸過程的能量需求。但是Hesselman[7]等人認為此比值直接反映了生物量中聚磷酸鹽量和糖原的含量。由于EBPR生物群落的不同,此比值并不能總是精確的符合模型提出的數值,而是變化的。
2.2以非醋酸鹽作為底物時EBPR的代謝作用
在實際的污水處理過程中,進水中可能不是僅僅含有醋酸鹽或短鏈脂肪酸,還可能存在其他種類的有機底物。因此,有必要研究以非醋酸鹽作為底物時EBPR的代謝作用。有關有機物是直接對EBPR起作用還是先轉化為揮發性脂肪酸再起作用這一點,還不是很清楚。Randall[8]等人認為揮發性脂肪酸的化學性質直接決定厭氧合成的PHA的數量和化學組成,進而決定好氧吸磷量。
Cech[9,10]等人經試驗證實以葡萄糖作為唯一碳源時也能實現EBPR。Carucci[11], Nakamura[12], Sudiana[13], Jeon[14]和Wang[15]等人經過試驗研究,提出并支持以葡萄糖作為碳源的EBPR的生物化學模型,而且還證實以葡萄糖作為碳源能實現穩定的EBPR。例如,C13核磁共振跟蹤數據表明在厭氧區乳酸菌吸收葡萄糖生成乳酸,最終以糖原的形式貯存在體內。Jeon[14] 等人描述的模型假定PAO在厭氧時將糖原轉化成PHA,而乳酸菌利用糖原合成聚乳酸鹽。在此模型中,PAO可以好氧吸磷。遺憾的是,除了對生物量中能觀察到的優勢細菌的有限的描述外,Jeon[14] 等人沒有證明實際的生物群落結構是否與模型提出的結構一致。Wang[15]等人提出了一個能解釋以葡萄糖作為碳源時的EBPR的不同模型。此模型的機理與Mino和Comeau-Wentzel提出的機理非常相似,但是他們也沒有提出直接的微生物學或生物化學的證據來支持該模型。
既然廢水中通常都存在氨基酸,那么氨基酸是否可以作為底物實現EBPR呢?Satoh[16,17]等人在試驗過程中發現谷氨酸鹽與天冬氨酸鹽都能作為唯一的碳源實現EBPR,他們認為厭氧時底物被貯存,但并不是所有被用作底物的谷氨酸鹽都以PHA的形式貯存在細胞內,相反,在貯存過程中合成了多肽,有關這一點還有待于進一步研究。
3.結語
有關EBPR的生物化學原理,研究者提出了許多模型。但是目前的生物化學模型仍不能解釋由試驗研究得出的所有試驗數據,例如微生物厭氧吸收底物時既沒有水解聚磷酸鹽也沒有將糖原轉變成PHA;在以葡萄糖作為碳源時也能實現EBPR,但在厭氧釋磷過程卻沒有形成PHA。這些結果反映出我們對EBPR的生物化學原理的認識是有限的。
隨著水體富營養化越來越嚴重,如何有效去除水中的營養物已經成為一個亟待解決的問題,而EBPR工藝由于其具有較高的除磷效率,因此在除磷方面將發揮重要的作用。為建立穩定可靠的EBPR系統,深入理解EBPR的生物化學原理是必需的,因此EBPR的生物化學原理是一個很有價值的研究課題。
參考文獻
[1] Comeau, H., Hall, K.J., Hancock, R.E.W. and Oldham, W.K.(1986)Biological model for enhanced biological phosphorus removal. Water Res. 20, 1511-1521.
[2] Wentzel, M.C., Lotter, L.H., Ekama, G.A., Loewenthal, T.E. and Marais, G.v.R. (1991)Evaluation of biochemical models for biological excess phosphorus removal. Water Sci. Technol. 23, 567^576.
[3] Mino, T., van Loosdrecht, M.C.M. and Heijnen, J.J.(1998)Microbiology and biochemistry of the enhanced biological phosphate removal process. Water Res. 32, 3193^3207.
[4] Van Loosdrecht, M.C.M., Hooijmans, C.M., Brdjanovic, D. and Heijnen, J.J. (1997) Biological phosphorus removal processes. Appl. Microbiol. Biotechnol. 48, 289^296.
[5] Kortstee, G.J.J., Appeldoorn, K.J., Bonting, C.F.C., van Niel, E.W.J.and van Veen, H.J.(2000)Ecological aspects of biological phosphorus removal in activated sludge systems. Adv. Microb. Ecol. 16, 169^ 200.
[6] Pereira, H., Lemos, P.C., Reis, M.A.M., Crespo, J.P.S.G., Carrondo, M.J.T. and Santos, H.(1996)Model for carbon metabolism in biological phosphorus removal processes based on in vivo 13C-NMR labeling experiments. Water Res. 30,2128^2138.
[7] Hesselmann, R.P.X., von Rummel, R., Resnick, S.M., Hany, R. and Zehnder, A.J.B.(2000)Anaerobic metabolism of bacteria performing enhanced biological phosphate removal. Water Res. 34, 3487^3494.
[8] Randall, A.A. and Liu, Y.-H. (2002) Polyhydroxyalkanoates form potentially a key aspect of aerobic phosphorus uptake in enhanced biological phosphorus removal. Water Res. 36, 3473^3478.
[9] Cech, J.S. and Hartman, P. (1990) Glucose induced breakdown of enhanced biological phosphate removal. Environ. Technol. 11, 651^656.
[10] Cech, J.S. and Hartman, P. (1993) Competition between polyphosphate and polysaccharide accumulating bacteria in enhanced biological phosphate removal systems. Water Res. 27, 1219^1225.
[11] Carucci, A., Majone, M., Ramadori, R. and Rossetti, S. (1994) Dynamics of phosphorus and organic substrates in anaerobic and aerobic phases of a sequencing batch reactor. Water Sci. Technol. 30, 237^246.
[12] Nakamura, K. and Dazai, M. (1986) Growth characteristics of batchcultured activated sludge and its phosphate elimination capacity. J. Ferment. Technol. 64, 433^439.
[13] Sudiana, I.M., Mino, T., Satoh, H., Nakamura, K. and Matsuo, T.(1999) Metabolism of enhanced biological phosphorus removal and non-enhanced biological phosphorus removal sludge with acetate and glucose as carbon source. Water Sci. Technol. 39, 29^35.
[14] Jeon, C.O. and Park, J.M. (2000) Enhanced biological phosphorous removal in a sequencing batch reactor fed with glucose as a sole carbon source. Water Res. 34, 2160^2170.
[15] Wang, N., Peng, J. and Hill, G. (2002) Biochemical model of glucose induced enhanced biological phosphorus removal under anaerobic condition. Water Res. 36, 49^58.
[16] Satoh, H., Mino, T. and Matsuo, T. (1990) Biochemical mechanism of anaerobic glutamate uptake in the enhanced biological phosphorus removal process. In: Proc. IAWPRC Biennial Conf. Kyoto, pp. 531^534.
[17] Satoh, H., Mino, T. and Matsuo, T. (1998) Anaerobic uptake of glutamate and aspartate by enhanced biological phosphorus removal activated sludge. Water Sci. Technol. 37, 579^582.
論文搜索
月熱點論文
論文投稿
很多時候您的文章總是無緣變成鉛字。研究做到關鍵時,試驗有了起色時,是不是想和同行探討一下,工作中有了心得,您是不是很想與人分享,那么不要只是默默工作了,寫下來吧!投稿時,請以附件形式發至 paper@h2o-china.com ,請注明論文投稿。一旦采用,我們會為您增加100枚金幣。



