高鐵酸鹽預氧化對藻類細胞的破壞作用及其助凝機理
劉偉 馬軍
摘 要 利用電子顯微鏡和紫外分光光度計檢測發(fā)現(xiàn),高鐵酸鹽對水中藻類細胞的表面結構有很大影響,破壞了細胞的表面鞘套。高鐵酸鹽的氧化作用導致的藻類細胞的鞘套破裂使其向水中釋放出胞內物質,這些生物高聚物能夠在混凝過程中起到助凝劑的作用。高鐵酸鹽分解后產生的氫氧化鐵膠體沉淀在藻類細胞的表面,也降低了藻類細胞的穩(wěn)定性。
關鍵詞 高鐵酸鹽;氧化;藻類;胞內物質;助凝
Effects of ferrate (Ⅵ) preoxidation on the architecture of algae cells and the mechanism of enhanced coagulation
LIU Wei, MA Jun
Abstract The scanning electron microscopy (SEM) and the ultraviolet spectro-photometer were used to measure the impact of ferrate (Ⅵ) preoxidation on the architecture of algae cells and the release of soluble organic matters from the cells. The results showed that ferrate (Ⅵ) preoxidation induced a number of clearly discernible effects on cell architecture, caused severe damage of cell layer. An excessive release of intracellular components into the surrounding media was also observed, which is possibly caused by ferrate stimulation or cleaved sheath by its oxidation. These biopolymers secreted in response to ferrate oxidation behave as a coagulant aid in coagulation process. Fe(OH)3 colloids derived from the decomposition of ferrate (Ⅵ) may precipitate on the surface of algae cells and thus decrease the stability of algae cells.
Keywords Ferrate (Ⅵ); oxidation; algae; intracellular organic matter; coagulant aid
湖泊與水庫是常用的地表水水源,其中含有大量的浮游生物。近年來,由于水體污染嚴重,在河流中也出現(xiàn)了藻類含量升高的情況。含藻水混凝困難,不易形成良好的絮體,沉淀效果差。水中的一些藻類能引起嗅味,影響飲用水的感官性狀;一些藻類會堵塞濾池,還有的藻類由于形體細小,可能會穿過濾池,造成濾后水的水質惡化。
前期的研究結果表明,高鐵酸鹽預氧化能夠有效地去除水中的藻類細胞[1],高鐵酸鹽預氧化可以大幅度降低腐植酸對藻類混凝去除的阻礙作用[2]。試驗中通過鏡檢發(fā)現(xiàn),水中的藻類細胞經(jīng)過高鐵酸鹽氧化后發(fā)生了結構和形態(tài)的變化,如藻類細胞表面被破壞,發(fā)生一定程度的聚集。在試驗中我們采用的原水藻類含量較高[1],約為8.28´106-2.26´107個/升,高鐵酸鹽投量則相對較低,為0.14-1.40mg/L [以(Fe)計],因此高鐵酸鹽預氧化后藻類去除率的大幅度提高不可能是單純氧化滅活作用所致。高鐵酸鹽對藻類具有氧化滅活作用的同時還可能存在著有利于后續(xù)混凝的作用。為進一步探討高鐵酸鹽氧化除藻的機理,我們采用電子掃描顯微鏡考察藻類細胞經(jīng)過高鐵酸鹽氧化后同正常細胞相比是否發(fā)生形態(tài)及結構變化,并測定處理前后水樣的紫外吸光度和高錳酸鹽指數(shù),考察水樣中溶解性有機物的變化規(guī)律。
1. 試驗過程和方法
1.1 藻類培養(yǎng)
為了消除水中其它因素對高鐵酸鹽預氧化除藻效果的影響,試驗所用藻類為人工培養(yǎng)。取含有夏季地表水中常見的綠球藻(Chlorococoam)和柵列藻(Scenedesmus)的湖水進行分級培養(yǎng)。首先配制適合兩種藻類生長的培養(yǎng)液,在小容積的玻璃容器中培養(yǎng)一段時間,在其藻類細胞濃度較高時轉移至大容器中繼續(xù)培養(yǎng)。無機培養(yǎng)液的濃度見表1,培養(yǎng)液中同時加入少量的土壤滲出液以刺激藻類的生長。培養(yǎng)溫度為15±1℃,起始pH 7.3,采用白熾燈和日光燈持續(xù)照明。每隔一天時間向容器中通入含1%CO2的混合空氣,以保證藻類光合作用的需要。在25天的培養(yǎng)后,培養(yǎng)液中的藻類細胞濃度為3.5-4.2´108個/升,pH 9.1,濁度20-40NTU。
表1 藻類培養(yǎng)液中各種無機鹽類的濃度
無機鹽
KNO3
Ca(NO3)2
MgSO4·7H2O
KH2PO4
FeCl3
濃度 (mg/L)
20
60
20
20
0.2
注:培養(yǎng)液中混合有0.7%的土壤滲出液。其制備方法是取1kg土壤,加入1L水煮沸60min,冷卻后沉淀,過濾后即為制得的土壤滲出液。
1.2 電子顯微鏡觀察
將未處理水樣(原培養(yǎng)含藻水)和處理后水樣(原水中加入高鐵酸鹽5mg/L氧化30min)各取一滴在干燥臺上,干燥2h。然后用Eiko IB-3型電子發(fā)射器向干燥后的水樣上鍍150nm的黃金。用Hitachi S-520型電子掃描顯微鏡觀察,加速電壓為15KV,并將藻類細胞顯微照相以進行對比。
1.3 燒杯攪拌試驗
靜態(tài)試驗在DBJ-621型六聯(lián)定時攪拌器上進行。將含藻水轉移至5個500 mL的燒杯中,用0.1N的HCI將原水的pH調至7.1。投加一定量的高鐵酸鹽 [以K2FeO4計],以200r/min的轉速攪拌一定時間,再投加一定量的混凝劑 [以Al2(SO4)3·18H2O計],繼續(xù)以200r/min的轉速攪拌1min,然后以40r/min的轉速攪拌10min,靜置20min,然后測定水樣的UV254吸光度和高錳酸鹽指數(shù)。高鐵酸鹽固體由改進的化學氧化法制備,主要成分為高鐵酸鉀。由于制備過程中加入了無機穩(wěn)定劑,該藥劑可以穩(wěn)定保存6-8個月,在水溶液中可以保持10h不分解,滿足使用要求。使用前配制成1g/L的使用液,根據(jù)前期試驗結果選擇投量5mg/L。
取一系列500mL水樣,進行單純高鐵酸鹽氧化(氧化時間1h)、單純硫酸鋁混凝、高鐵酸鹽預氧化(氧化時間1h)后用硫酸鋁混凝等不同的靜態(tài)攪拌試驗(攪拌試驗工藝同上),處理后水樣經(jīng)過0.45 mm的濾膜過濾,然后在200 nm-320 nm波長范圍內進行紫外掃描。
2. 試驗結果與討論
2.1 電子顯微鏡掃描結果
顯微鏡中觀察到培養(yǎng)液背景純凈,藻種比較單一,主要為綠球藻和柵列藻。典型的柵列藻(Scenedesmus)如圖1a所示:每個單體具有4個細胞,在外側的兩個細胞的尾端并生著兩根長刺。4個細胞尺寸幾乎相同。柵列藻的整個細胞體被鞘套包裹(這是指圖中的網(wǎng)狀外層)[3],可以清楚地看到細胞表層上的瘤狀突起,這也是柵列藻的典型表層結構。每個細胞表面沿縱軸方向的條痕也清晰可見。綠球藻(Chlorococoum)活體細胞呈橢圓形,細胞體外對生的長刺有規(guī)律地分布在細胞周圍(圖1b)。圖中的兩種藻類細胞與在鏡檢過程中觀察到的活體細胞相比有一點收縮,這是由于掃描電鏡檢測的前處理(干燥過程)造成的。
由高鐵酸鹽氧化前后藻體細胞的電子掃描顯微鏡對比結果可以看出,高鐵酸鹽預氧化對藻體細胞結構造成了明顯的影響。高鐵酸鹽氧化處理促使藻體細胞向周圍的介質中釋放了大量的胞內物質(圖2a, 圖3a)。這種現(xiàn)象可能是由于高鐵酸鹽刺激了藻體細胞(圖3a),造成其過度分泌胞內物質;或者是氧化破壞了細胞的鞘套(圖2a)使其外殼開裂,從而造成藻類細胞胞內物質的外流。
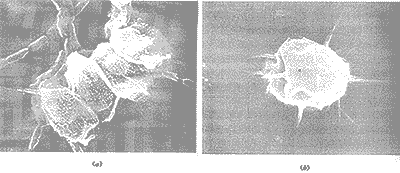
(a)四細胞的柵列藻,尾端細胞長有長刺;(b)綠球藻周圍規(guī)律地排列對生長刺。放大倍數(shù):(a) ×4000; (b) ×8000。
圖1 未處理的藻類細胞
高鐵酸鹽氧化對藻類細胞的另一個破壞作用就是造成了外部鞘套的強烈卷繞。顯微照片的(圖2b, 圖3a)測定結果表明,高鐵酸鹽預氧化使細胞表面結構受到嚴重破壞,原來柵列藻(Scenedesmus)的瘤狀突起排列消失,外部鞘套受到破壞;而綠球藻(Chlorococoum)在氧化后細胞體塌陷,外部的長刺脫落。

a)釋放胞內物質;(b)外部鞘套卷繞;(c)Fe(OH)3膠體沉淀在藻細胞表面。放大倍數(shù):(a) ×4000; (b) ×4000; (c) ×6000。
圖2 高鐵酸鹽預氧化處理后的柵列藻細胞
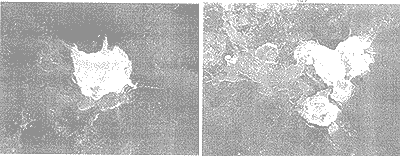
(a)釋放胞內物質;(b)細胞的聚集。放大倍數(shù):(a) ×7000; (b) ×4000。
圖3 高鐵酸鹽預氧化處理后的綠球藻
上述高鐵酸鹽氧化對藻細胞的兩種影響可能同時存在,即高鐵酸鹽氧化首先造成藻細胞體外殼的卷繞,并刺激其過度分泌胞內物質,進一步的氧化作用會使細胞鞘套開裂致使胞內物質大量外流。
高鐵酸鹽預氧化的特點是分解后會形成氫氧化鐵膠體沉淀 [Fe(OH)3]。圖2c顯示氫氧化鐵膠體沉淀到藻類細胞表面的情況,這些沉淀顯然會改變藻類的表面性質,如藻細胞的表面電性。一旦這些沉淀物吸附到藻細胞的表面,它們就降低了藻細胞的活動性,而且增加了藻細胞的沉淀性。高鐵酸鹽預氧化后部分藻細胞發(fā)生聚集(圖3b)。從顯微攝影結果可以看出,藻細胞釋放的胞內物質使它們發(fā)生聚集,F(xiàn)e(OH)3沉淀的絮凝作用也會促進這種聚集作用,或者是這兩者協(xié)同作用結果使藻類細胞在混凝之前就發(fā)生初步的凝聚,從而易于被硫酸鋁混凝沉淀。
從電子顯微鏡的掃描照片可以看出,高鐵酸鹽預氧化給藻類細胞的表面結構帶來了一系列影響,破壞了藻細胞外部鞘套,造成細胞內物質向周圍介質的釋放,并改變其沉降性能。藻類的混凝、沉淀是基于硫酸鋁水解產物的電中和原理,除了藻細胞的表面電性,水中的溶解性有機物也是影響混凝效果的重要水質因素。高鐵酸鹽氧化造成的藻類細胞的胞內物質釋放必然使水中的有機物濃度或者數(shù)量發(fā)生變化。
2.2 水中溶解性有機物的變化
將原水和處理過的水樣經(jīng)0.45mm的醋酸纖維膜過濾后在200nm-320nm波長范圍內紫外掃描,測定紫外吸光度(UVA)。紫外的吸光度(254nm波長)是水中天然有機物(NOM)的良好替代參數(shù)。在水處理中,波長254nm處的吸光度值被用于近似地反映水的溶解性有機碳(DOC)[4]。紫外掃描結果可以反映水中天然有機物的芳香化程度。在試驗中UVA和UV254用于指示處理前后水樣的溶解性有機物(DOM)的濃度變化。
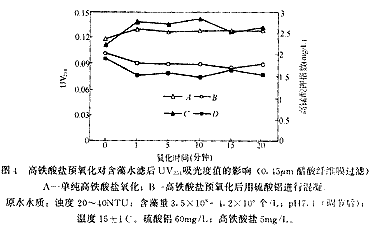
圖4是在不同氧化時間下,經(jīng)過和不經(jīng)過硫酸鋁混凝處理含藻水UV254吸光度和高錳酸鹽指數(shù)變化情況,時間0點為不投加高鐵酸鹽的情況。可以看出,含藻水的UV254吸光度值在很短的氧化時間內有較大的升高,但隨著氧化時間的延長吸光度變化不大。高鐵酸鹽預氧化后用硫酸鋁混凝沉淀后UV254值低于單純硫酸鋁混凝,同樣隨時間的延長變化不大。高錳酸鹽指數(shù)也表現(xiàn)出同樣的變化規(guī)律。一些研究者在使用氧化劑從污水中去除藻類時發(fā)現(xiàn)溶解性有機碳(DOC)升高的現(xiàn)象[5-6],使用O3、ClO2和Cl2等處理含藻水也會使DOC及UV254升高[7]。高鐵酸鹽預氧化使水樣UV254和高錳酸鹽指數(shù)升高說明氧化作用可能增加了含藻水中有機物含量或者數(shù)量,經(jīng)過硫酸鋁混凝后這些指標下降說明混凝過程可以去除這些增加量,并且低于單純硫酸鋁混凝。
從圖中曲線的形狀可以看出,短時間(1分鐘)內的氧化就使UV254和高錳酸鉀指數(shù)發(fā)生較大變化,說明高鐵酸鹽的氧化造成藻細胞釋放胞內物質的過程在很短的時間內就可以完成。試驗中高鐵酸鹽的特征紫色在10分鐘內才完全褪去,這說明高鐵酸鹽的部分氧化性被水中的其他還原性物質(如有機物)所消耗,高鐵酸鹽可能會氧化藻細胞釋放的胞內物質。
A-單純高鐵酸鹽氧化;B-高鐵酸鹽預氧化后用硫酸鋁進行混凝。原水水質:濁度 20-40NTU;含藻量3.5×108-4.2×108 個/L;pH 7.1(調節(jié)后);溫度15±1℃。硫酸鋁 60mg/L;高鐵酸鹽5mg/L。
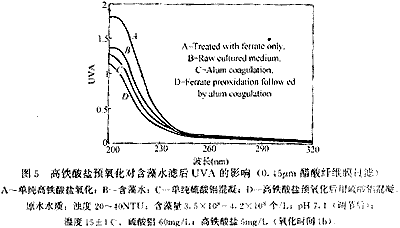
與圖4相同,在波長為200nm-320nm之間的紫外掃描值也表現(xiàn)出同樣的變化規(guī)律(圖5)。經(jīng)過高鐵酸鹽氧化后水樣的紫外吸光度值整體升高,尤其是在200nm-250nm波長范圍內升高的幅度較大。硫酸鋁混凝處理后UVA有一定程度的下降,但是幅度不大。高鐵酸鹽預氧化后用硫酸鋁混凝則使UVA顯著下降,低于單純硫酸鋁混凝。多數(shù)有機物在紫外區(qū)間都有吸收,紫外吸光度值曲線的整體升高說明高鐵酸鹽氧化后藻類細胞可能釋放出多種有機物而不是一種物質。一般來說,200nm-250nm波長范圍內的強吸收帶可能是由于存在雙鍵并處于共軛狀態(tài),高鐵酸鹽氧化可能使水中帶有雙鍵基團(如羰基等)的有機物濃度或數(shù)量增加,這些有機物可能是來自藻類細胞釋放也可能是高鐵酸鹽氧化有機物后生成,它們可能會被后續(xù)的混凝去除,因而高鐵酸鹽預氧化后用硫酸鋁混凝水中的溶解性有機物(DOM)進一步降低。
Herth[8-10]報道幾種綠藻能夠向水中釋放出幾丁質纖維,另外藻類細胞還可能釋放出除幾丁質之外的其他有機物質。這些綠藻或者藍綠藻釋放出的胞內物質在水中會具有帶負電荷或中性多聚電解質的性質,并因此而具有一定的絮凝作用。
高鐵酸鹽預氧化對水中藻類細胞的作用機理可能是:高鐵酸鹽預氧化能夠破壞藻細胞的表面結構,造成藻細胞表面鞘套的卷繞,并可能使細胞的外鞘開裂,致使胞內物質外流。高鐵酸鹽分解后產生的氫氧化鐵膠體也可以被吸附在一些藻類細胞表面,在降低細胞的表面電荷的同時也增加了這些細胞的沉淀性。氫氧化鐵膠體的吸附和胞內物質的絮凝作用在混凝之前就能夠使部分藻類細胞發(fā)生凝聚,胞內物質在混凝過程中還可能進一步起到助凝劑的作用。這是高鐵酸鹽預氧化具有優(yōu)良除藻效果的重要原因。
A-單純高鐵酸鹽氧化;B-含藻水;C-單純硫酸鋁混凝;D-高鐵酸鹽預氧化后用硫酸鋁混凝。原水水質:濁度 20-40NTU;含藻量3.5×108-4.2×108 個/L;pH 7.1(調節(jié)后);溫度15±1℃。硫酸鋁 60mg/L;高鐵酸鹽5mg/L(氧化時間1h)。
3. 結論
電子顯微鏡的觀察結果表明,高鐵酸鹽預氧化能夠改變水中藻類細胞的表面性質,破壞細胞表面結構,并刺激胞體向其周圍的介質中釋放出胞內物質。紫外掃描測定結果表明,高鐵酸鹽預氧化能增加水中有機物濃度。這些胞內物質具有大分子結構,在混凝過程中可能起到助凝劑的作用。另外,高鐵酸鹽分解后產生的Fe(OH)3膠體能夠吸附并沉淀到藻細胞的表面,從而降低了藻細胞的穩(wěn)定性。
參考文獻
[1] 馬軍,石穎,劉偉,李圭白. 高鐵酸鹽復合藥劑預氧化除藻效能研究[J]. 中國給水排水,1998, 14 (5):9~11.
[2] 馬軍,劉偉,盛力,李圭白. 腐植酸對高鐵酸鉀預氧化除藻效果的影響[J]. 中國給水排水,2000, 16 (9):5~8.
[3] Starhlin L. A. and Pickett-Heaps J. D. The ultra-structure of Scenedesmus (chlorophydeae). Ⅰ. Species with the “reticulate” or “wary” type of ornamented layer[J]. J. Phycol., 1975, (11): 163-185.
[4] Owen D. M., Amy G. L., Chowdhury Z. K., Paode R., McCoy G. and Viscosil K. NOM characterization and treatability[J]. J. AWWA, 1995, (87): 46-63.
[5] Betzer N. and Kott Y. Effect of halogens on algae[J]. Wat. Res. 1969, (3): 237-264.
[6] Echelberger W. F., Pavoni J. L., Singer P. C. and Tenny M. W. Disinfections of algae-laden waters[J]. J. sanit. Engng Div. Am. Soc. Civ. Engrs 1971, (172): 721-730.
[7] Sukenik A., Teltch B., Wachs A. W., Shelef G., Nir I. and Levanon D. Effect of oxidants on microalgal flocculation[J]. Wat. Res. 1987, 21(5): 533-539.
[8] Herth W. A special chitin-fibril-synthesizing apparatus in centric diatom[J]. Cyclotella. Naturwissenschaften, 1978, (65): 260-269.
[9] Herth W. The site of b-chitin fibril formation in centric diatomsⅡ. The chitin-forming cytoplasmic structures[J]. J Ultrastru. Res., 1979, (68): 16-27.
[10] Herth W., Bartholtt W. The site of b-chitin fibril formation in centric diatom Ⅰ.Pores and fibril formation[J]. J Ultrastru. Res., 1979, (68): 6-15.
論文搜索
月熱點論文
論文投稿
很多時候您的文章總是無緣變成鉛字。研究做到關鍵時,試驗有了起色時,是不是想和同行探討一下,工作中有了心得,您是不是很想與人分享,那么不要只是默默工作了,寫下來吧!投稿時,請以附件形式發(fā)至 paper@h2o-china.com ,請注明論文投稿。一旦采用,我們會為您增加100枚金幣。



